{kind=link}
{kind=link}
{kind=link}

I am a post-doctoral fellow in the Takahashi Laboratory. My main interest is in auditory representation of sound location and sound structure in the auditory system. We work on the barn owl, Tyto alba, an accomplished nocturnal hunter.
The barn owl, which is renowned for its ability to locate prey by hearing alone, possesses a topographically organized map of auditory space. This space-map allows easy access to spatial sound stimuli. When sounds move in the real world, there is a resultant and predictable change in activity in the space map. Precisely how does the owl use this auditory space map? To answer questions about neuronal function, one must be able to first develop a reliable method of assessing behavior. Measuring neuronal function is most useful when it has a real, measurable impact.
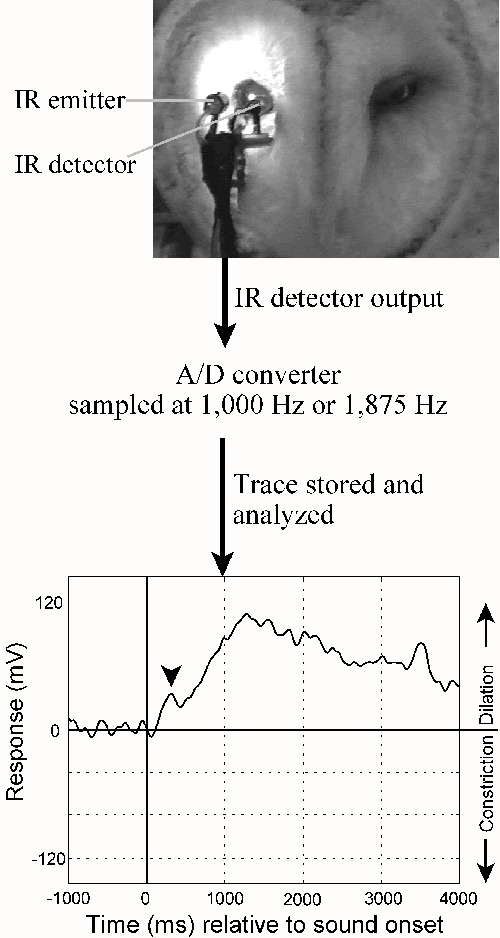
I found that when a sound stimulus is played in a quiet room, the barn owl responds by dilating its pupil. This reflexive sound is not particular to the owl: most vertebrates, including humans, exhibit this reflex. It has formed the basis of popular myths such as the ability to tell truths from lies by looking at the pupil (perhaps possible, but extremely unreliable so far). It was best exemplified in the truth detectors used in the cult movie Blade Runner, and the current science fiction serial Stargate SG-1. This dilation of the pupil is easily measured in the owl as well as in humans, and forms the basis of my work. While the pupil dilates to the first presentation of a sound stimulus, the response decreases if the stimulus is repeated. This process is called habituation. It is easy to imagine how a change in environment first evokes a big response, and as it becomes commonplace, the subject becomes inured to the sound, making it a part of the altered environment. Thus, after a few weeks of combat, soldiers may sleep through gunfire, and yet be woken by an alarm clock. In this case, the alarm clock is the 'unusual' stimulus, while the gunfire is what they have gotten accustomed to. The process by which a habituated subject responds to an unusual or novel stimulus is called 'recovery'.
Why do we habituate to a repeated sound? So that we can ignore common sounds, and retain our sensitivity to fresh changes in our surrounds. While the owl habituates to repeated presentations of a given sound, the presentation of any new sound stimulus again evokes a pupillary dilation response. I found that the magnitude of this recovered response was directly proportional to how different the ‘novel’ sound was from the ‘habituating’ sound. This acoustically induced pupillary dilation response (PDR), was described a research paper in 2000 (Bala and Takahashi, J.Comp.Physiol.A, 186, 425-34). I habituated my owl subjects to repeated presentations of sound from a given location. Then I presented sounds from incrementally increasing angular separations. Upto a point, the subject was not able to detect the movement of the sound source. However, when the sounds were sufficiently separated, the owl could detect the change, and we could measure that detection behavior by a recovered dilation response. The magnitude of this recovered response was stimulus dependent: when the location of the novel stimulus was furthest from the location of the habituating stimulus, the magnitude of the recovery was greatest. Using this method, I was able to determine that the smallest discriminable angular separation in the owl – minimum audible angle – was 3°.
Next, we compared behavior, measured using the PDR, with responses from neurons that constitute the owl’s topographic auditory space map. Neurons that make up the space map are themselves ‘spatially tuned’, responding to sounds from a small region of space, but not to sounds from other regions of space. Thus, when sounds are moved in space, the response of the neurons changes. We can ask, by moving sounds across small regions of space, how easily barn owl auditory neurons detect changes in spatial location. Our results, described in a recent Nature article (Bala, Spitzer, Takahashi; Nature 424, 771-774, 2003), showed that the performance of individual neurons could match or exceed behavioral performance. This put to rest any necessity to invoke computational models that help boost the performance of auditory neurons, models which have previously been postulated. In addition, behavioral threshold was very close to the mean threshold of the neuronal population: this indicated that the ‘lower-envelope principle’, which predicts that behavior is related to the performance of the most sensitive neurons, is not applicable to the barn owl’s discrimination of auditory space. Thus the answer was that behavior was determined instead by contributions from the whole neuronal population.
Currently, I am trying to understand the impact of noisy signals on discrimination, as opposed to
signals that are very consistent from trial to trial. I am also exploring the activity of single
nerve cells in the awake owl, and how that activity changes during behavioral trials.