![]() Note: Conversion to HTML for this document is in
progress!
Note: Conversion to HTML for this document is in
progress!
Reprinted as:
University of Oregon
Institute of Cognitive & Decision Sciences
Technical Report No. 93-3
Copyright (c) 1992, 1995, Jennifer J. Freyd
We showed subjects a line-drawn creature moving on a computer screen. Suddenly the creature stopped and subjects' memory for position was tested, revealing large forward memory shifts. Representational momentum was reduced when the creature moved tail-first instead of head-first, corresponding to a subjective experience of watching an impaired or sick animal. An "inanimate" icon condition produced no directionality effect. We discuss the significance of dynamics, directionality, and animacy in event and object perception.
Prof. Jennifer J. Freyd Office: (503) 346-4929 Department of Psychology Lab: (503) 346-4950 University of Oregon FAX: (503) 346-4911 Eugene, OR 97403 Internet: jjf@dynamic.uoregon.edu
[Slide 1: Sculpture Photo -- Pavlova and Mordkin][1]
In everyday encounters with the world we interact with a dynamic environment.
[Slide 2: Painting Reproduction -- Religious Scene]
Many events involve motor action from the perceiver that is related to action by others.
[Slide 3: Wildlife Photo -- Mountain Goats]
Even if the current environment contains no active objects besides the perceiver, the perceiver may well be planning to act on the environment, by walking through it or by manipulating an object within it.
[Slide 4: Wildlife Photo -- Snake]
Or the perceiver may be monitoring the environment for possible and probable events, such as the possible sudden lunge of a resting animal. The advantages to anticipating movement suggest to me that representations of the world emphasize future time (Freyd, 1987, 1992, 1993). Even static stimuli can invoke dynamic mental representations.
[Slide 5: Wildlife Photo -- Deer Running Down Hill]
I have reported before (Freyd, 1983, 1987) that subjects misremember some static stimuli -- such as frozen-action photographs -- as further along the path of frozen motion. In a related series of studies, Ron Finke and I (e.g.: Freyd & Finke, 1984, 1985; Freyd, 1992)demonstrated that when a rotation of a visual pattern is implied, an observer's memory for the pattern orientation tends to be displaced forward in the direction of the implied rotation. We called this representational momentum.
 Slide 6: RM Task & Shift, Freyd
& Finke, 1985
Slide 6: RM Task & Shift, Freyd
& Finke, 1985
[VIDEO -- F&F part]
In one study (Freyd & Finke, 1985) subjects were presented with a
static figure in a sequence of orientations sampled from a possible
path of rotation. Subjects were instructed to remember the third
orientation they saw, and were presented with a fourth orientation
that was either the same as, or different from, the third. Test
orientations were varied parametrically around True-Same. We found a
generally symmetric unimodal distribution of "same" responses
centered not on true-same but on a forward rotation from true-same.
That is, subjects showed a shift in memory for position.
 Slide 7: Velocity, Freyd &
Finke, 1985
Slide 7: Velocity, Freyd &
Finke, 1985
As with physical momentum, representational momentum is proportional to the implied velocity of motion (Freyd & Finke, 1985).
 Slide 8: Retention Interval, Freyd
& Johnson, 1987
Slide 8: Retention Interval, Freyd
& Johnson, 1987
In addition, the amount of memory distortion follows a continuous stopping function for the first 250 ms or so of the retention interval (Freyd & Johnson, 1987). It also is impervious to feedback or practice (Finke & Freyd, 1985).
These parametric effects have been demonstrated in a non-visual domain: Using sequences of pitches, Mike Kelly, Mike DeKay, and I (Freyd, Kelly, DeKay, 1990) replicated the basic phenomenon and showed that it behaved similarly to the visual case with changes in implied velocity, implied acceleration, and retention interval.
[Slide 9: Wildlife Photo -- Monkeys]
In more recent work my colleagues and I have been applying the concept of dynamic representations to questions about event and object perception. We have been influenced by the work of a number investigators, including Hubbard, Bharucha, Cooper, Faust, Verfaillie, d'Ydewalle, and Ranney, who have contributed to the representational momentum literature (e.g.: Cooper, Gibson, Mawafy, & Tataryn, 1987; Faust, 1990; Hubbard, 1990; Hubbard & Bharucha, 1988; Ranney, 1989; Verfaillie & d'Ydewalle, 1991). Much of this work has demonstrated that representational momentum is affected by various aspects of object and event perception.
[Slide 10: Blurry Street Scene]
In our laboratory in Oregon, we are using the representational momentum paradigm to probe the nature of event perception. Currently, for instance, Amy Hayes and I are using representational momentum to investigate the role of object and event coherence, and Donna McKeown and I are studying the role of dynamic representations in contributing to aesthetic excitement when viewing static art (Freyd, 1993; McKeown & Freyd, 1992).
[Slide 11: Wildlife Photo -- Wolves]
Today I will report on two experiments from a series of studies being completed in collaboration with Geoffrey Miller, who is currently enjoying a postdoctoral fellowship in Sussex England.
[Slide 12: Wildlife Photo -- Eagles]
Miller and I are interested in the relationship between dynamic representations and animate motion perception. We considered the mounting evidence that humans are exquisitely sensitive perceivers of biological motion (also Miller & Freyd, 1993). For instance, Maggie Shiffrar and I reported that with sufficient processing time apparent motion paths are consistent with anatomically possible motion even if that motion is not the simplest, or shortest, rigid displacement (Freyd & Shiffrar, 1988; Shiffrar & Freyd, 1990, 1991).
David Perrett and his colleagues have identified numerous cells in the macaque superior temporal sulcus which are conjointly selective for particular combinations of biological forms and movements (Perrett, Smith, Mistlin, Chitty, Head, Potter, Broennimann, Milner, & Jeeves, 1985; Perrett, Harries, Mistlin, & Chitty, 1990). These motion sensitive cells respond vigorously to moving primate figures and are unresponsive to moving inanimate control objects
[Slide 13: Wildlife Photo -- Baby Leopard]
These results, and evolutionary logic -- Miller's specialty -- suggested that dynamic representations might be tuned toward laws of animate motion.
[Slide 14: Wildlife Photo -- Llama]
Miller and I considered the role of directionality in animate motion. We hypothesize that perceivers use various cues such as the position of sensory organs and the aerodynamic structure of animate creatures to anticipate motion in certain directions.
[Slide 15: Wildlife Photo -- Horses]
The majority of animals with which humans interact move nose-first and eye-first as opposed to tail-first.
 Slide 16: Wildlife Photo --
Dolphins[2]
Slide 16: Wildlife Photo --
Dolphins[2]
Many animals are also pointy, or streamlined, in the direction of frequent body motion. Miller and I hypothesized that these attributes of animals would be used by perceivers in anticipatory dynamic representations.
We predicted that representational momentum shifts would thus be greater for forward than backward moving creatures.
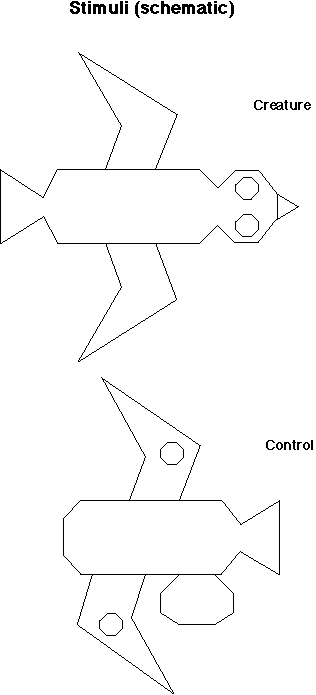 Slide
17: Schematic Creature Stimuli
Slide
17: Schematic Creature Stimuli
In one experiment we used a line drawing of a bird-like creature moving on a computer screen either head-first or tail-first. We also had an inanimate control icon.
 Slide
18: Design for Creature RM
Slide
18: Design for Creature RM
32 Subjects participated.
[VIDEO: Creature Sections]
The video monitor shows the forward creature block. The filming of the display exaggerated the brightness of the head and adds some jerkiness to the motion as compared with the computer display.
In each trial an icon traverses an Oval Path 2 Times. Then the icon stops and disappears. Next is a retention interval of 250 msec; then a test icon appears.
The subject indicates "Same" or "Different"
We had one between subject factor, Rotation Direction (Clockwise vs Counterclockwise). We had two within subject blocked factors,
Animacy (the Creature vs Control icon) and Facing Direction (Forward vs Backward Motion). And we had two within subject factors randomized within each block: Starting/Stopping Position (top vs bottom of oval path) and Test Position (-3, -2, -1, 0, 1, 2, 3).
We predicted a main effect for Representational Momentum, a main effect for Animacy, and an Interaction between Animacy and Facing Direction
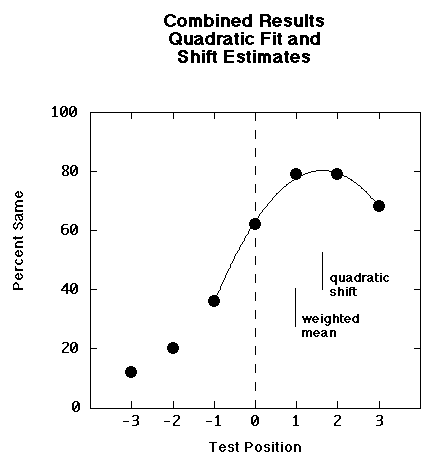 Slide
19: Quadratic Fit for Creature RM
Slide
19: Quadratic Fit for Creature RM
This slide shows the results collapsed across all subjects and all conditions except test position. We found a robust forward memory shift. The peak of the shift was well captured by a quadratic function.
We used the distribution of same responses by test position to derive a measure of central tendency. This slide shows the estimate of memory shift using the quadratic fit. It also shows a more conservative estimate of memory shift based on all the data points, the weighted mean. For subsequent analyses we will use weighted means calculated for each subject and stimulus condition.
 Slide 20: Anova for Creature RM
Slide 20: Anova for Creature RM
For each subject and condition we calculated the weighted mean. A 5-way Anova revealed: a Main effect for Representational Momentum, a Main effect for Facing, an Interaction between Facing and Animacy, and an Interaction between Rotate and Stopping Position. No other main effects or interactions were found. (The Rotate/Stopping Interaction reflects a rightward bias; this bias does not interact with other factors.)
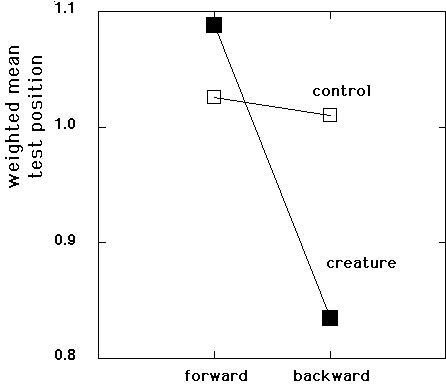 Slide
21: Interaction, Creature RM
Slide
21: Interaction, Creature RM
The interaction between facing and animacy is displayed in this slide. The difference between forward and backward motion is highly significant for the creature conditions.
In our next experiment we investigated subjective experience of these stimuli.
 Slide 22: Design Creature
Survey
Slide 22: Design Creature
Survey
24 Subjects watched the same stimulus displays but without the memory task. After each of the 4 experimental blocks subjects completed a questionnaire with 13 questions.
 Slide
23: Questionnaire Page 1
Slide
23: Questionnaire Page 1
Subjects were instructed: "We are interested in your perceptions and impressions of the moving object that is being displayed. For each question below, please indicate your answer by placing an 'X' on the scale. . . .None of the questions below have right or wrong answers. Some questions may seem metaphorical, speculative, or silly. Just try to give us your impressions as accurately as possible."
In general, high scores would indicate subjective experience of a fast, active, and well-functioning object, low scores, a slow, passive, and impaired object. For example, Question 1 was: "How fast does the object appear to be moving?" Subjects selected from very slow to very fast. Questions 5: "Does the object appear to be active or passive?" Subjects selected from passive to active. And Question 6 : "Does the object appear to be functioning well or not?" Subjects selected from not function to functioning.
 Slide 24: Creature Survey Anova
Slide 24: Creature Survey Anova
We then measured the position of the x mark from the left side of the line. An anova revealed main effects for all factors we tested and all but one interaction. Of most interest to us was the significant main effect for animacy.
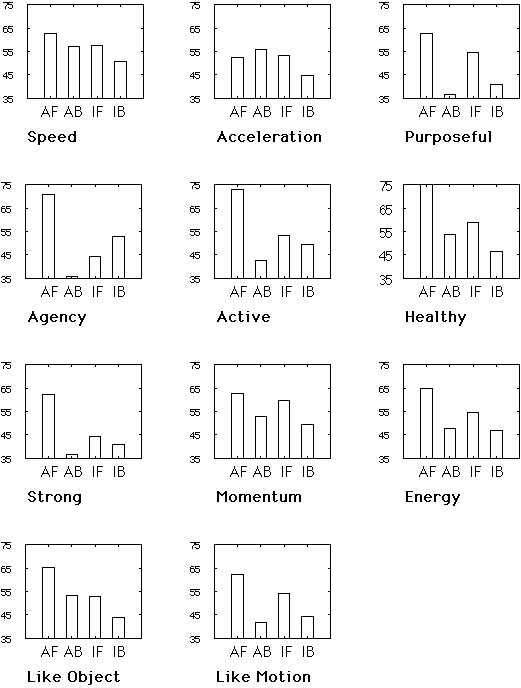 Slide
25: Creature Survey Individual Graphs
Slide
25: Creature Survey Individual Graphs
This slide shows results for the 11 individual questions for which we predicted that forward, animate motion would produce the largest responses. In each histogram, the first bar is for the Forward Animate or Creature motion, the second bar is for Backward Animate motion, and the remaining bars show the results for the Inanimate Control Icon. Interestingly, some of the most marked effects were for questions assessing the subjective qualities of healthy versus sick functioning and active versus passive motion.
This corresponded to subjects written responses. For the forward creature they wrote: "Looks like a bird looking for bugs to eat." "It looks like a bird of prey, circling an animal, and preparing to dive." For backward creature they wrote: "Backward flying bird." "I think that object seems like a bee, and I don't like bees." And for the control object: "It is very odd in form. Not aerodynamic at all, but it seems to be flying." "It looks a lot like a boomerang and a milk bottle."
What do these results tell us about animacy, directionality and dynamic representations? The subjective experience scale results show large effects of animacy and directionality. However, the representational momentum results really only show a directionality effect, not a demonstrable animacy effect.
 Slide 26: Consistent and
Inconsistent Arrow Stimuli
Slide 26: Consistent and
Inconsistent Arrow Stimuli
In fact, in earlier work I completed with Teresa Pantzer, we demonstrated a directionality effect for an inanimate icon (Freyd, 1989; Freyd & Pantzer, in preparation). We used an arrow in a representational momentum experiment. We compared cases where the implied motion was consistent with the arrow directionality with cases where the motion was inconsistent.
 Slide 27: Arrow Results
Slide 27: Arrow Results
We found that the consistent motion led to larger representational momentum shifts than did the inconsistent motion.
[Slide 28: Ambiguous Figure -- Duck/Rabbit]
Perhaps representational momentum is influenced by directionality, but not by object identity per se. Alternatively, perhaps our stimuli, moving around as if with internal force, whether creature or control icons, activated mechanisms that evolved to track and anticipate animate motion. If so, directionality might be subordinate to animacy. Perhaps only neuroanatomic evidence will settle this issue.
[Slide 29: Wildlife Photo -- Giraffe]
In the meantime, I conclude with the thought that this research demonstrates that event perception and event memory is influenced by anticipation of future action. More specifically, event perception and memory are influenced by anticipation of the future direction of action. This is as it should be for clever, planning big-brained creatures ... such as ourselves.
Cooper, L. A., Gibson, B. S., Mawafy, L., & Tataryn, D. J. (1987). Mental extrapolation of perceptually-driven spatial transformations. In Twenty-eighth Annual Meeting of the Psychonomic Society, November, 1987. Seattle, WA: Psychonomic Society.
Faust, M. (1990). Representational momentum: A dual process perspective. Doctoral dissertation, University of Oregon.
Finke, R. A., & Freyd, J. J. (1985). Transformation of visual memory induced by implied motions of pattern elements. Journal of Experimental Psychology: Learning, Memory, and Cognition, 11, 780-794.
Freyd, J. J. (1983). The mental representation of movement when static stimuli are viewed. Perception & Psychophysics, 33, 575-581.
Freyd, J. J. (1987). Dynamic mental representations. Psychological Review, 94, 427-438.
Freyd, J. J. (1989). Arrows and triangles moving in the mind. Short paper given before the Inaugural Fred Attneave Lecture, University of Oregon, May 1, 1989.
Freyd, J. J. (1992). Dynamic representations guiding adaptive behavior. In F. Macar, V. Pouthas, &. J. Friedman (Eds.), Time, Action and Cognition: Towards Bridging the Gap (pp. 309-323). Dordrecht: Kluwer Academic Publishers.
Freyd, J. J. (1993). Five hunches about perceptual processes and dynamic representations. In D. E. Meyer & S. Kornblum (Ed.), Attention and Performance XIV: Synergies in Experimental Psychology, Artificial Intelligence, and Cognitive Neuroscience -- A Silver Jubilee. Cambridge, MA: MIT Press.
Freyd, J. J. & Finke, R. A. (1984). Representational momentum. Journal of Experimental Psychology: Learning, Memory, and Cognition, 10, 126-132.
Freyd, J. J. & Finke, R. A. (1985). A velocity effect for representational momentum. Bulletin of the Psychonomic Society, 23, 443-446.
Freyd, J. J. & Johnson, J. Q. (1987). Probing the time course of representational momentum. Journal of Experimental Psychology: Learning, Memory, and Cognition, 13, 259-268.
Freyd, J. J., Kelly, M. H., & DeKay, M. (1990). Representational momentum in memory for pitch. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 1107-1117.
Freyd, J. J. & Pantzer, T. M. (in preparation). Static patterns moving in the mind. In S. M. Smith, T. B. Ward, & R. A. Finke (Ed.), The Creative Cognition Approach. Cambridge, MA: MIT Press
Freyd, J. J. & Shiffrar, M. (1988) Violations of shortest-path constraints in apparent motion using full-bodied stimuli. Paper given at the Twenty-ninth Annual Meeting of the Psychonomic Society, Chicago, Illinois, November 10-12, 1988.
Hubbard, T. L. & Bharucha, J. J. (1988). Judged displacement in apparent vertical and horizontal motion. Perception & Psychophysics, 44, 211-221.
Hubbard, T. L. (1990). Cognitive representations of linear motion: possible direction and gravity effects in judged displacements. Memory & Cognition, 18, 299-309.
McKeown, D. & Freyd, J.J. (1992). Dynamic aspects of static art. Poster presented at the 1992 Convention of the American Psychological Society, San Diego, June 1992
Miller, G. & Freyd, J.J. (1993). Dynamic mental representations of animate motion: The interplay among evolutionary, cognitive & behavioral dynamics. Technical Report: University of Sussex at Brighton CSRP 290, ISSN 1350-3162.
Perrett, D., Harries, M., Mistlin, A.J., & Chitty, A.J. (1990). Three stages in the classification of body movements by visual neurons. In Images and Understanding, (Eds.) Barlow, H.B., Blakemore, C. & Weston-Smith, M. Cambridge University Press, 94-107.
Perrett, D., Smith, P., Mistlin, A., Chitty, A., Head, A., Potter, D. Broennimann, R., Milner, A., & Jeeves, M. (1985). Visual Analysis of body movements by neurones in the temporal cortex of the macaque monkey: A preliminary report. Behavioural Brain Research, 16, 153-170.
Ranney, M. (1989). Internally represented forces may be cognitively penetrable: Comment on Freyd, Pantzer, and Cheng. Journal of Experimental Psychology: General, 118, 399-402.
Shiffrar, M., & Freyd, J. J. (1990). Apparent motion of the Human body. Psychological Science, 1, 257-264.
Shiffrar, M., & Freyd, J. J. (1991). Where did the time go? Temporal factors in apparent motion path choice. In Thirty-second Annual Meeting of the Psychonomic Society, . San Francisco, California: Psychonomic Society.
Verfaillie, K., & d'Ydewalle, G. (1991). Representational momentum and event course anticipation in the perception of implied periodical motions. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 302-313.
The research reported here was supported by NSF Presidential Young Investigator Award BNS-8796324 and NIMH Grants R01-MH39784 and K02-MH00780 to J.J. Freyd.